Partenogénesis en reptiles
Parthenogenesis in reptiles
Resumen
La mayor parte de los organismos eucariontes son gonocóricos y se reproducen sexualmente, sin embargo, en algunos grupos han surgido linajes que están constituidos por un solo sexo y se reproducen clonalmente. Entre los reptiles del orden Squamata existen alrededor de 40 especies de lagartijas que se reproducen mediante partenogénesis, que es un mecanismo de reproducción clonal a través del cual las hembras producen descendientes genéticamente idénticos a ellas. La partenogénesis obligada es un proceso reconocido ampliamente en los reptiles, pero en años recientes se ha documentado la partenogénesis facultativa en hembras de diferentes especies de lagartijas y serpientes que han sido criadas en cautiverio.
Palabras clave: Meiosis, partenogénesis, reptiles, Squamata
Abstract
The greater part of eukaryotes are gonochoristic and reproduce sexually, nevertheless, in some groups have arisen lineages constituted by only one sex that reproduce clonally. Among reptiles of the order Squamata there are about 40 species of lizards that reproduce by parthenogenesis that is a mechanism of clonal reproduction through which, females produce offspring genetically identical to them. Obligate parthenogenesis is a process widely recognized in reptiles, but recently has been documented facultative parthenogenesis in females of different species of lizards and snakes, that have been kept in captivity.
Keywords: Meiosis, parthenogenesis, reptiles, Squamata
Introducción
La reproducción sexual es el tipo de reproducción predominante entre los eucariontes, sin embargo, en varios linajes se han originado especies que están constituidas por un solo sexo. De acuerdo con diversos autores, una de cada mil especies multicelulares de eucariontes es unisexual (Simon et al., 2003). Estas especies se originan a partir de ancestros con ambos sexos (gonocóricos) y presentan mecanismos de reproducción en los que la meiosis (división celular que da origen a los gametos) está alterada. Gracias a esto se inhibe la recombinación genética y se producen especies con descendientes genéticamente idénticos a sus progenitores, a través de alguno de los tipos de reproducción clonal (Avise, 2008).
La reproducción clonal se presenta en varios grupos de eucariontes, principalmente en plantas, rotíferos, nematodos, artrópodos y en aproximadamente 100 especies de vertebrados, entre los que se encuentran peces, anfibios y reptiles. En estos últimos, la reproducción clonal es exclusiva de algunas especies del orden Squamata (Vrijenhoek et al., 1989).
Partenogénesis y origen de las especies partenogenéticas de reptiles
Diversos procesos citológicos que inhiben o alteran la meiosis son los responsables de la existencia de distintos tipos de reproducción clonal entre los eucariontes, pero sólo uno de ellos (partenogénesis) está presente en las especies unisexuales de reptiles (Cuellar, 1971). La partenogénesis obligada o telitoquia se caracteriza porque los gametos se producen sin reducción en la ploidia y se desarrollan en ausencia de espermatozoides, en organismos genéticamente idénticos a la madre (Figura 1). Lo anterior conduce a la formación de especies constituidas exclusivamente por hembras (los machos no existen), en las que la meiosis está presente y el número de cromosomas es restituido gracias a la fusión de los gametos con los cuerpos polares (originados en la primera o segunda división meiótica) o a la endoduplicación (Cuellar, 1971; 1987).
La endoduplicación o endomitosis es un proceso citológico que consiste en una mitosis premeiótica sin cariocinesis, la cual da como resultado la duplicación del número cromosómico en las células pregaméticas (Cuellar, 1971). Dichas células presentan pares de cromosomas idénticos y entran a un proceso de meiosis que da lugar a la producción de gametos con el número de cromosomas propio de la especie. Varios autores han sugerido que la endomitosis es responsable de la restitución en varias especies unisexuales de vertebrados, sin embargo, la meiosis de estos organismos ha sido poco estudiada, a pesar de ser un proceso clave en la generación de la reproducción clonal (Simon, 2003).
Por otra parte, diversos estudios han mostrado que los linajes unisexuales de lacertilios se han originado espontáneamente o por hibridación (Figura 2). La pérdida espontánea de la reproducción sexual puede ocurrir en hembras inicialmente diploides y gonocóricas, por medio de mutaciones en los genes involucrados en el proceso meiótico. Dichas mutaciones provocan la aparición de hembras que producen gametos no reducidos, capaces de dar origen a nuevos linajes unisexuales (Cuellar 1987). El origen espontáneo de la partenogénesis es un proceso documentado ampliamente en invertebrados, pero entre los vertebrados clonales únicamente ha sido encontrado en lacertilios del género Lepidophyma: L. reticulatum y L. flavimaculatum. Esta última especie presenta poblaciones gonocóricas (México y norte de Centroamérica) y partenogenéticas (Costa Rica y Panamá) que son morfológica y genéticamente homogéneas (Sinclair et al., 2009).
La hibridación interespecífica es otro de los mecanismos relevantes en el origen de especies unisexuales de lacertilios (Avise et al., 1992), varios autores consideran que la producción de gametos no reducidos y la capacidad de reproducirse clonalmente son resultado directo de la alteración que la combinación de genomas distintos produce en el proceso meiótico de los híbridos. El origen híbrido ha sido comprobado en una gran cantidad de especies unisexuales y parece ser el mecanismo de origen más importante en vertebrados, ya que prácticamente todas las especies clonales de peces, anfibios y reptiles se han producido gracias al apareamiento entre dos especies distintas (Cuellar 1987).
La hibridación inicial entre especies gonocóricas conduce a la formación de linajes unisexuales diploides; sin embrago, más del 60% de los vertebrados unisexuales de origen híbrido son poliploides (Vrijenhoek et al., 1989). De acuerdo con varios autores, las especies triploides se originan cuando se lleva a cabo la fecundación de los gametos diploides de una hembra unisexual por parte de los machos de sus especies parentales o por machos de una especie distinta.
En el surgimiento de las especies poliploides hay un punto de coincidencia entre las dos teorías, ya que los seguidores de la teoría del origen espontáneo sugieren que una especie triploide puede originarse una vez que una hembra que adquirió la capacidad de reproducirse clonalmente se cruza con un macho de una especie distinta (Figura 2; Cuellar, 1987).
El origen híbrido de varias especies unisexuales de vertebrados fue sugerido con base en estudios morfológicos, ecológicos, cariológicos y aloenzimáticos, pero en años recientes el uso de marcadores moleculares del DNA nuclear (nDNA) y mitocondrial (mtDNA) ha permitido esclarecer varios aspectos sobre el origen de las especies clonales (Avise et al., 1992). Los marcadores nucleares de herencia codominante (con expresión de alelos provenientes de ambos padres) en combinación con el mtDNA (de herencia materna) han permitido confirmar el origen por hibridación de varias especies y han revelado la dirección del cruzamiento original (Avise, 1994; 2000).
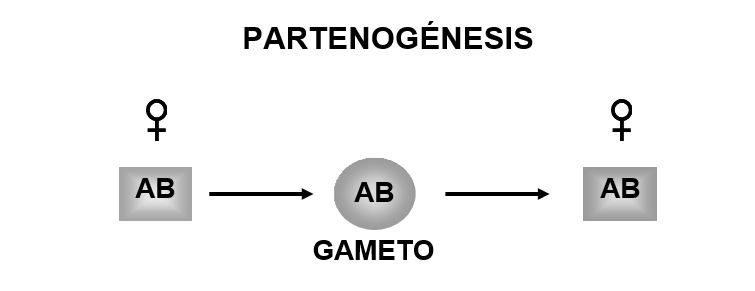
Figura 1.
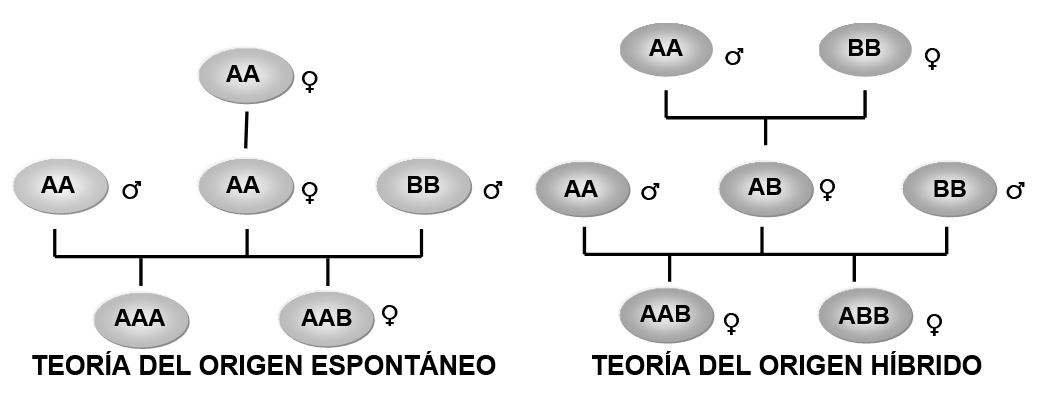
Figura 2.
Diversidad clonal
Aunque las especies unisexuales se reproducen clonalmente, diversos estudios han mostrado que la mayor parte de ellas presentan cierto grado de diversidad genética (MacCulloch et al., 1995; Fu et al., 1999). La variabilidad genética presente en las especies unisexuales (diversidad clonal) es producida por recombinación (durante la meiosis) o mutación, posteriores al origen de la partenogénesis (Parker et al., 1989). Estudios realizados en varios animales unisexuales revelan que estos organismos presentan una sensibilidad alta a los factores ambientales y variación fenotípica de magnitud similar a la que presentan las especies gonocóricas (Lynch y Gabriel, 1983).
En las especies partenogenéticas la principal fuente de variación es la mutación de tipo postformacional y ha sido evaluada con base en estudios de morfología, cariología, histocompatibilidad, aloenzimas y DNA (Manríquez Morán, 2007). Los estudios realizados a la fecha, muestran que las especies unisexuales de lagartijas presentan poca variación a nivel morfológico (Parker y Selander, 1984), sin embargo, existen varias especies con distribuciones amplias que están compuestas por clones que presentan diferencias en tamaño y patrones de coloración y escutelación (Parker et al., 1989; Walker et al., 1997; Hernández Gallegos et al, 1998; Manríquez Morán, 2002). Dentro de estas últimas se encuentran los teiidos Aspidoscelis tesselata y A. maslini, que son especies constituidas por varios clones geográficamente aislados que difieren en sus patrones de coloración (Manríquez Morán 2002). Un patrón similar se presenta cuando las especies partenogenéticas se analizan cariológicamente, pues la mayor parte de ellas presentan cariotipos híbridos altamente heteromórficos (con dos complementos haploides diferentes), pero con poca variación intraespecífica. Sin embargo, algunas especies que muestran distribuciones amplias y ocupan hábitats heterogéneos pueden presentar algunas variaciones en su cariotipo (Fritts, 1969; Lowe et al., 1970; Cole 1979; Manríquez-Morán, 2002). Por ejemplo, Aspidoscelis maslini, A. sonorae y A. exsanguis presentan cariotipos que parecen haberse originado gracias a la modificación del cariotipo híbrido inicial, por medio de transformaciones como las fusiones y principalmente, las fisiones cromosómicas.
Sin embargo, la caracterización genética de las especies partenogenéticas es lo que ha permitido detectar con mayor certeza su diversidad genética y establecer el número real de clones que las están formando. La estructura genética de las especies unisexuales de vertebrados fue establecida de manera indirecta con base en los resultados proporcionados por los estudios de histocompatibilidad realizados con transplantes de piel. Dichos estudios han permitido identificar a especies partenogenéticas constituidas por un solo clon (A. tesselata, A. neomexicana, A. velox, A. maslini, A. rodecki, A. laredoensis, A. cozumela; Maslin, 1967; Cuellar, 1976; Cordes et al., 1990; Cuellar y Wright, 1992; Hernández Gallegos et al., 1998; Abuhteba et al., 2000; Hernández Gallegos et al., 2003; Manríquez Morán y Méndez de la Cruz, 2008) o bien, a complejos de especies integrados por clones de origen y ploidía distintos.
En décadas recientes los marcadores moleculares (nucleares y mitocondriales) han sido los que han permitido establecer, de manera más confiable, la variabilidad existente dentro de las especies con reproducción clonal. Las aloenzimas mostraron que varias de las especies unisexuales del género Aspidoscelis están integradas por clones geográfica y genéticamente distinguibles que se han generado gracias a mutaciones posteriores al origen de la reproducción clonal (Dessauer y Cole, 1989; Manríquez-Morán, 2007). Lo anterior es congruente con los estudios en los que se ha utilizado el DNA, ya que los análisis realizados con enzimas de restricción o secuenciación han mostrado que varias de las especies unisexuales formalmente reconocidas están constituidas por clones más inclusivos y genéticamente diferentes (Moritz et al., 1989; Manríquez-Morán, 2002). Una excepción a esto la presentan varias de las especies de lagartijas unisexuales del género Darevskia, ya que sus mtDNA´s son constantes a lo largo de su distribución e, incluso, son compartidos por varias especies unisexuales (Murphy et al., 2000).
La variabilidad genética en las especies partenogenéticas depende de factores filogenéticos y ecológicos, así como del tiempo de origen de los taxones, por lo que se considera que existe una correlación entre la diversidad de este tipo de organismos y su área de distribución, los hábitats que ocupan y su edad. Se ha visto que las especies unisexuales que han invadido áreas extensas y heterogéneas tienen un origen antiguo y están integradas por dos o más clones con características genéticas distintas (Manríquez Morán, 2002).
Hábitats de las especies partenogenéticas
El origen de la partenogénesis en lacertilios puede ser azaroso, pero una vez que surge una hembra capaz de reproducirse clonalmente, su éxito y el surgimiento de una especie nueva depende de la interacción de los individuos unisexuales con sus especies parentales. Cuando se encuentran en simpatría, la competencia puede evitar su establecimiento, pero diversos trabajos han mostrado que las especies clonales suelen presentarse en sitios que son desfavorables para sus especies parentales. Recientemente se ha documentado que una gran cantidad de especies partenogenéticas, tanto de invertebrados como de vertebrados, habitan en islas o sitios que se comportan como islas (Cuellar, 1994). Son comunes en playas marinas y lacustres, cuerpos de agua y zonas abiertas dentro de las selvas, troncos caídos, hormigueros y termiteros, cuevas, etc.
Partenogénesis facultativa
Dentro de los escamados, la partenogénesis obligada fue descubierta por el herpetólogo ruso Ilia Darevsky hace casi 56 años, sin embargo, en años recientes se ha documentado la existencia de partenogénesis facultativa. Este tipo de reproducción se ha observado en hembras de diversas especies de lagartijas y serpientes (Varanus komodoensis, V. panoptes, V. ornatus, Python molurus, Thamnophis elegans, T. marcianus, Crotalus horridus, C. unicolor y Acrochordus arafurae), que han permanecido en cautiverio y en ausencia de machos. A diferencia de lo que ocurre en la partenogénesis obligada, las hembras que se reproducen facultativamente suelen dar origen a descendientes machos. Lo anterior se debe a la condición homóciga de los individuos, ya que en los escamados en los que se ha presentado este tipo de partenogénesis las hembras son heterogaméticas (ZW), mientras que todas las crías producidas presentan una combinación ZZ. Sin embargo, en el pitón P. morulus se comprobó la producción de hembras genéticamente idénticas a la madre (Booth et al., 2012).
En algunos casos se ha observado que las hembras que se reproducen facultativamente posteriormente pueden interactuar con machos y producen crías por reproducción sexual.
La partenogénesis facultativa no ha sido registrada en poblaciones silvestres, pero se cree que pudo haberse originado como un mecanismo alternativo para momentos en los que no se tiene acceso a una pareja, o bien, como una forma de modificar la proporción de sexos en alguna población (Watt et al., 2006).
Independientemente del tipo de partenogénesis (obligada o facultativa), la meiosis es el proceso clave para la existencia de este tipo de reproducción. Sin embargo, el conocimiento que se tiene de este proceso y su modificación en las especies partenogenéticas es prácticamente nulo. Por lo anterior, su estudio y entendimiento es necesario para explicar las diferencias entre los distintos linajes con reproducción clonal.
Referencias
Avise, J. C. (1994). Molecular markers, natural history and evolution. Chapman y Hall, New York, U. S. A.
Avise, J. C. (2008). Clonality. The genetics, ecology, and evolution of sexual abstinence in vertebrate animals. Oxford University Press, New York, U. S. A.
Avise, J. C., Quatro, J. M., & Vrijenhoek, R C. (1992). Molecular clones within organismal clones: Mitochondrial DNA phylogenies and the evolutionary histories of unisexual vertebrates. Evolutionary Biology, 26:225-246.
Booth, W., Smith, C. F., Eskridge, P. H., Hoss, S. K., Mendelson, J. R., & Schuett, J. W. (2012). Facultative parthenogenesis discovered in wild vertebrates. Biology Letters, 53:671-688.
Cole, C. J. (1979). Chromosome inheritance in parthenogenetic lizards and evolution of allopolyploidy in reptiles. The Journal of Heredity, 70:95-102.
Cuellar, O. (1971). Reproduction and the mechanism of meiotic restitution in the parthenogenetic lizard Cnemidophorus uniparens. The Journal of Morphology, 133: 139-166.
Cuellar, O. (1987). The evolution of parthenogenesis: A historical perspective. En P. B. Moens (ed.), Meiosis. Academic Press, New York , pp. 43-103
Cuellar, O. (1994). Biogeography of parthenogenetic animals. Biogeographica, 70:1-13.
Dawley, R. M. (1989). Evolution and ecology of unisexual vertebrates. En R. M. Dawley y J. P. Bogart (eds.), An introduction to the unisexual vertebrates. New York State Museum Bulletin 466, New York, pp. 1-18.
Fritts, T. H. (1969). The systematics of the parthenogenetic lizards of the Cnemidophorus cozumela complex. Copeia, 1969:519-535.
Fu, J., Murphy, R. W., & Darevsky, I. S. (1999). Limited genetic variation in Lacerta mixta and its parthenogenetic daughter species: Evidence from cytochrome b and ATPase 6 gene DNA sequences. Genetica, 105:227–231.
Hernández Gallegos, O., Manríquez Morán, N. L., Méndez de la Cruz, F. R., Villagrán Santa Cruz, M. & Cuellar, O. (1998). Histocompatibility in parthenogenetic lizards of the Cnemidophorus cozumela complex from the Yucatan Peninsula of México. Biogeographica, 74:117-124.
Lowe C. H., Wright, J. W., Cole, C. J., & Bezy, R. L. (1970). Natural hybridization between the teiid lizards Cnemidophorus sonorae (parthenogenetic) and Cnemidophorus tigris (bisexual). Systematic Zoology, 19:114-127.
Lynch, M., & Gabriel, W. (1983). Phenotypic evolution and parthenogenesis. The American Naturalist, 122:745-764.
MacCulloch, R. D., Murphy, R. W., Kupriyanova, L. A., Darevsky, I. S., & Danielyan, F. D. (1995). Clonal variation in the parthenogenetic rock lizard Lacerta armeniaca. Genome, 38:1057–1060.
Manríquez Morán, N. L. (2002). Origen y diversidad clonal en las especies de lagartijas partenogenéticas del complejo Cnemidophorus cozumela (Reptilia: Teiidae). Tesis de Doctorado, Instituto de Biología, Universidad Nacional Autónoma de México. México.
Manríquez Morán, N. L. (2007). Diversidad clonal en los lacertilios unisexuales del género Aspidoscelis. Boletín de la Sociedad Herpetológica Mexicana, 15:1-12.
Manríquez Morán N. L., & Méndez de la Cruz, F. R. (2008). Genetic homogeneity between two populations of the parthenogenetic lizard Aspidoscelis cozumela. Revista Mexicana de Biodiversidad, 79:421-426.
Maslin, T. P. (1968). Taxonomic problems in parthenogenetic vertebrates. Systematic Zoology, 17:219-231.
Parker, E. D., & Selander, R. K. (1984). Low clonal diversity in the parthenogenetic lizard Cnemidophorus neomexicanus (Sauria: Teiidae). Herpetologica, 40:245-252.
Parker, E. D., Walker, J. M., & Paulissen, M. A. (1989). Clonal diversity in Cnemidophorus: ecological and morphological consequences. En R. M. Dawley y J. P. Bogart (eds.), Evolution and ecology of unisexual vertebrates. New York State Museum Bulletin 466, New York, pp. 72–86.
Simon, J. C., Delmotte, F., Rispe, C., & Crease, T. (2003). Phylogenetic relationships between parthenogens and their sexual relatives: the possible routes to parthenogenesis in animals. Biological Journal of the Linnean Society, 79:151-163.
Sinclair, E. A., Pramuk, J. B., Bezy, R. L., Crandall, K. A., & Sites, J. W. (2010). DNA evidence for non hybrid origins of parthenogenesis in natural populations of vertebrates. Evolution, 64:1346-1357.
Vrijenhoek, R. C., Dawley, R. M., Cole, C. J., & Bogart, J. P. (1989). A list of known unisexual vertebrates. En R. M. Dawley y J. P. Bogart (eds.), Evolution and ecology of unisexual vertebrates. New York State Museum Bulletin 466, New York, , pp. 19–23.
Watt. P. C., Buley, K. R., Sanderson, S., Boardman, W., Ciofi , C., & Gibson, R. (2006). Parthenogenesis in Komodo dragons. Nature, 444:1021-1022.
Zweifel, R. G. (1965). Variation in and distribution of the unisexual lizard, Cnemidophorus tesselatus. American Museum Novitates, 2235:1-49.
[a] Profesora-Investigadora del Área Académica de Biología de la Universidad Autónoma del Estado de Hidalgo. Desarrolla las líneas de investigación de sistemática molecular y reproducción clonal en reptiles.